RUI: Collaborative Research: Microbiological Survey and Inventory of Gregarines Parasitizing Aquatic and Riparian Insects of
the Texas Big Thicket
R. E. Clopton, Principal Investigator
NSF Award ID: 0340774 (05/2004-04/2008)
Project Goals and Objectives
The project surveys aquatic and riparian insect communities and their gregarine parasites across 9 ecosystems of the Texas Big Thicket. We describe and document all taxa; develop new identification documents; distribute comprehensive ecological, community, and specimen databases; disseminate optimized collection, preservation, and staining protocols; and, create a public specimen base to support future systematic revision of the Nearctic gregarines. The project contributes to our long-term goal of comprehensive discovery and systematic revision of the Nearctic Eugregarinida. The project has 6 major goals:
- Survey the insect and gregarine communities from the 9 Big Thicket ecosystems
- Establish an ecosystem correlated public specimen base of Big Thicket aquatic and riparian insects; and the first public gregarine specimen base for the southern Nearctic fauna
- Permanently document all insect and gregarine taxa collected with permanent specimens, description of all new taxa and redescription of known taxa as appropriate, new identification keys for gregarines and Big Thicket insects, and systematic revision with resolution of nomenclatural issues as appropriate
- Test new hypotheses exploring the effects of host insect metamorphosis, trophic habit, niche, and latitude on patterns of gregarine diversity
- WWW distribution of all project products
- Develop expertise in gregarine and insect taxonomy and systematics through a cross-institutional research and training collaboration including the full integration and mentoring of graduate and undergraduate students in all phases of the project
Research and Education Activities
Undergraduate training
We trained 13 undergraduate students and 2 masters student in field collection and field preservation of specimens, preparation of permanent specimens for museum deposition, digital image acquisition and analysis; taxonomic scholarship, identification, and analysis; and, molecular techniques and phylogeny reconstruction.
Field Collections and Biodiversity Discovery
Field Collections
We sampled all 9 traditional Big Thicket ecosystems (Beech-Magnolia-Loblolly Pine; Longleaf Pine Upland ; Oak-Gum Floodplain; Palmetto-Hardwood Flats; Slash Pine Monoculture; Pond, lake, and bog systems; 1st, 2nd, and 3rd order streams; Cypress-Tupelo; and, Pine Savanna Wetlands). Additionally we sampled odonates from 10 localities representing the 6 biotic provinces of Texas, 4 localities in Belize, Central America and 10 localities along a corridor extending from eastern Texas through Oklahoma, Kansas and Nebraska.
Insect survey
We collected 10,893 insects representing at least 9 orders,53 families, 136 genera, and 224 species. We prepared 5,285 insects as permanent museum specimens for deposition in the Sam Houston State University Insect Collection.
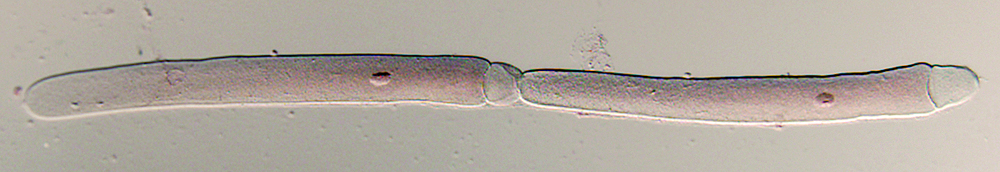
Gregarine survey
We processed 9,583 insects and other arthropods representing over 500 host morphotaxa across 4 arthropod classes, 15 orders, 42 families, and 64 putative genera. Of the 8,838 hosts examined in postmortem dissection, 4,210 were infected by at least 1 gregarine species (47.6% overall prevalence). We prepared 4,200 permanent slide preparations of gregarine parasites for deposition in the HWML, University of Nebraska State Museum, UNL, Lincoln, NE as well as 110 samples of gregarine tissue representing 49 gregarine morphotaxa for subsequent molecular analysis. With the support of the 2006 REU Supplement to REC, we developed a stable primer set and extraction/amplification protocol for gregarine 18s ribosomal sequence data. To date we have obtained complete 18s ribosomal sequence data for 34 gregarine taxa. Our sampling exceeds our initial host diversity sampling goals: we have sampled 157% of projected insect orders (11 of 7), 4 non-insect arthropod orders and 100% of projected host morphotaxa (500 of 500). Gregarine diversity is less rich but more unknown than we projected for a sample of this size: we have recovered 126 morphotaxa of which 101 are new putative species (80%). In our proposal we estimated that a sample of our current size would yield at least 182 morphotaxa, of which 72% would be new to science. Our expanded collecting efforts have and 7 putative new species and 2 putative new gregarine genera from odonates in Belize, Central America as well as confirming the expanded distribution of known gregarine genera parasitizing odonates from Nebraska to Belize.
Findings
Insect Survey
The insect survey results confirm the trends predicted in our project proposal. Most of the species we have collected are the first records from the counties surveyed and the Big Thicket region. This reflects the generally poor state of existing insect survey in the target region. The insect survey results indicate relatively high community diversity and species richness compared to most Nearctic surveys. Rather than a few significantly dominant species within aquatic communities in the Big Thicket, individuals are more evenly distributed among member taxa. We have not collected as many new insect species as we had anticipated, rather we appear to be sampling in a distributional intersection where the geographic boundaries of many Western and Eastern faunal species appear to converge in the Big Thicket. These samples present new opportunities to re-evaluate the uncertain systematic position and taxonomic status of several genera and species known only from the centroids of their putative distributions. In addition, we are collecting many taxa, especially from minor groups, that have not been reported from the Nearctic since the mid-twentieth century.
Gregarine Survey
Our collections and observations to date suggest that the diversity and faunal distribution of gregarine parasites in insect populations differs among Northern and Southern Nearctic communities. These differences may be functions of the timing and availability of exogenous energy sources, food chain length, or host population size constraints that arise as community diversity increases at lower latitudes. Northern host communities are less diverse, more redundant, and less even than comparable host communities sampled in this project. These differences in host community structure may underlie latitudinal differences in gregarine transmission and species richness. There is no significant latitudinal shift in richness of gregarine genera, at least among gregarine communities infecting damselflies. However, there do appear to be shifts in species richness among genera: in general, neotropical gregarine communities appear to be more redundant and subject to species dominance effects than their counterparts in the temperate zone. Molecular and phylogenetic analyses based on samples collected during the gregarine survey have allowed us to postulate a phylogeny of the septate gregarines and assess major ranks within the existing systematic. We have proposed new high-level morphological character and reviewed existing morphological characters as part of a larger family level revision of the septate gregarines.
Outreach Activities
I regularly give talks to public groups (Kiwanis and Optomist Clubs, the public nature centers, K-12 science classes, etc.) My talks tend to focus on biodiversity, the role of invertebrate diversity in human life, the importance of basic science in American society, and the role of NSF as a funding agency for basic science.
Permanent Collections
Insects
10,893 insects representing at least 9 orders,53 families, 136 genera, and 224 species. We prepared 5,285 insects as permanent museum specimens for deposition in the Sam Houston State University Insect Collection. The Sam Houston State University and Texas A&M University Insect Collections are active research collections with access and lending policies to support academic and public sector research and development.
Gregarines
4,200 permanent slide preparations of gregarine parasites were prepared for deposition in the HWML, University of Nebraska State Museum, UNL, Lincoln, NE. The H. W. Manter Laboratory for Parasitology, University of Nebraska State Museum, is an active research collection with access and lending policies to support academic and public sector research and development. Its holdings can be searched and loans requested via the WWW.
Publications
Cielocha, J. J., T. J. Cook, R. E. Clopton. 2011. Host Utilization and Distribution of Nubenocephalid Gregarines (Eugregarinorida: Actinocephalidae) Parasitizing Argia spp. (Odonata: Zygoptera) in the Central United States. Comparative Parasitology 78(1):152-160. PDF
Cook, T. J., R. E. Clopton, And J. L. Cook. 2011. Neoasterophora megasylvaticus n. gen., n. sp. (Apicomplexa: Eugregarinida: Actinocephalidae: Pileocephalinae) Parasitizing Cheumatopsyche analis (Trichoptera: Hydropsychidae) in the Texas Big Thicket. Comparative Parasitology 78(1):56-62. PDF
Clopton, R. E., T. J. Cook, and J. J. Cielocha. 2010. Nubenocephalus nickoli n. sp. and Nubenocephalus xunantunichensis n. sp. (Apicomplexa: Eugregarinida: Actinocephalidae) Parasitizing Damselflies (Odonata: Zygoptera) in Belize, Central America. Comparative Parasitology 77(2):125-136. PDF
Clopton, R. E. 2009. Phylogenetic Relationships, Evolution, and Systematic Revision of the Septate Gregarines (Apicomplexa: Eugregarinorida: Septatorina). Comparative Parasitology 76(2): 167-190. PDF
Clopton, R. E., T. J. Cook, and J. L. Cook. 2008. Gregarina tropica n. sp. (Apicomplexa: Eugregarinorida: Gregarinicae: Gregarinidae) Parasitizing the Brown-winged Earwig, Vostox brunneipennis (Dermaptera: Labiidae), in the Texas Big Thicket. Comparative Parasitology 75:215-227. PDF
Clopton, R. E., T. J. Cook, and J. L. Cook. 2008. Trichurispora wellgundis n. g., n. sp. (Apicomplexa: Eugregarinida: Hirmocystidae) parasitizing adult Water Scavenger Beetles, Tropisternus collaris (Coleoptera: Hydrophilidae) in the Texas Big Thicket. Comparative Parasitology 75: 82-91. PDF
Realzola, E., J. L. Cook, T. J. Cook, and R. E. Clopton. 2007. Composition of gyrinid aggregations in the East Texas Primitive Big Thicket. Coleopterist's Bulletin 61:495-502.
Hays, J. J., R. E. Clopton, T. J. Cook and J. L. Cook. 2007. Revision of the Genus Nubenocephalus and Description of Nubenocephalus secundus N. Sp. (Apicomplexa: Actinocephalidae) Parasitizing Adults of Argia sedula (Odonata: Zygoptera: Coenagrionidae) in the Primitive Texas Big Thicket, U.S.A. Comparative Parasitology. 74: 286-293. PDF
Clopton, R.E., T. J. Cook and J. L. Cook. 2007. Revision of Geneiorhynchus Schneider, 1875 (Apicomplexa: Eugregarinida: Actinocephalidae: Acanthosporinae) with Recognition of 4 New Species of Geneiorhynchus and Description of Geneiorhynchus manifestus N. Sp. Parasitizing Naiads of the Green Darner, Anax junius (Odonata: Aeshnidae) in the Texas Big Thicket. Comparative Parasitology 74: 273-285. PDF
Contributions
The project contributes to the discipline's on-going effort to document and understand the extent and pattern of biodiversity on Earth. We are focused on the biological diversity and association of a host-parasite system: namely the parasitic gregarines (Protista) and the insect hosts in the Big Thicket region of Texas. Current knowledge of biodiversity patterns indicates a distinct latitudinal effect on biodiversity: communities become richer (more species) as one approaches the equator. Richness is only one measure of diversity. Dominance and evenness (the relative distribution of individuals among species in a community) indicates deeper energy and trophic relationships within the community. Classically, each host species represents the opportunity for one or more parasite species to arise and associate by co-evolution and co-speciation, thus one expects parasite richness to track host richness. However, parasites depend upon energetic and trophic relationships within a community for transmission among hosts. Thus the relative dominance of species or the evenness of host population distribution across host richness may be a more important indicator of underlying parasite richness than simple host richness. This study samples a southern Nearctic region for both insect and gregarine diversity and compares the results to an existing survey conducted in Nebraska. We sampled all 9 traditional Big Thicket ecosystems and the contemporary slash pine monoculture ecosystem. Additionally we sampled odonates from 10 localities representing the 6 biotic provinces of Texas, 4 localities in Belize, Central America and 10 localities along a corridor extending from eastern Texas through Oklahoma, Kansas and Nebraska. In all, we collected 20,476 insects and other arthropods representing at least 4 arthropod classes, 15 orders,53 families, 136 genera, and 500 species. Of the 8,838 hosts examined in postmortem dissection, 4,210 were infected by at least 1 gregarine species (47.6% overall prevalence). Gregarine diversity is less rich but more unknown than we projected for a sample of this size: we have recovered 126 morphotaxa of which 101 are new putative species (80%). In our proposal we estimated that a sample of our current size would yield at least 182 morphotaxa, of which 72% would be new to science. Our expanded collecting efforts have and 7 putative new species and 2 putative new gregarine genera from odonates in Belize, Central America as well as confirming the expanded distribution of known gregarine genera parasitizing odonates from Nebraska to Belize. The insect survey results indicate relatively high community diversity and species richness compared to most Nearctic surveys. Rather than a few significantly dominant species within aquatic communities in the Big Thicket, individuals are more evenly distributed among member taxa. We appear to be sampling in a distributional intersection where the geographic boundaries of many Western and Eastern faunal species appear to converge in the Big Thicket. Our collections and observations to date suggest that the diversity and faunal distribution of gregarine parasites in insect populations differs among Northern and Southern Nearctic communities. These differences may be functions of the timing and availability of exogenous energy sources, food chain length, or host population size constraints that arise as community diversity increases at lower latitudes. Northern host communities are less diverse, more redundant, and less even than comparable host communities sampled in this project to date. These differences in host community structure may underlie latitudinal differences in gregarine transmission rates and thus gregarine species richness. There is no significant latitudinal shift in richness of gregarine genera, at least among gregarine communities infecting damselflies. However, there do appear to be shifts in species richness among genera: in general, neotropical gregarine communities appear to be more redundant and subject to species dominance effects than their counterparts in the temperate zone. Molecular and phylogenetic analyses based on samples collected during the gregarine survey have allowed us to postulate a phylogeny of the septate gregarines and assess major ranks within the existing systematic. We have proposed new cardinal morphological character and reviewed existing morphological characters as part of a larger revision of a major family level revision of the septate gregarines.

Collecting at Beaverslide Pond, Oak-Gum flood plain, Big Thicket National Preserve, Texas.

Water lillies, lake near Village Creek, Sandy Land Unit, Big Thicket National Preserve, Texas.

Longleaf Pine Upland, Sandy Land Unit, Big Thicket National Preserve, Texas.

First order stream in Beech-Magnolia-Loblolly Pine ecosystem, Big Thicket National Preserve, Texas.

Picking invertebrates, Big Thicket National Preserve, Texas.

Big Sandy Creek, Beech-Magnolia-Loblolly Pine ecosystem, Big Thicket National Preserve, Texas.

They don't call it the "Smoking Pistol" pit barbeque for nothing. Cold Springs, Texas.

Double-Lake, near Big Thicket National Preserve, Texas.

Oak-Gum Flood plain ecosystem, Big Thicket National Preserve, Texas.